Cell Res:复旦大学徐彦辉团队揭示了SNAPc结合的RNA聚合酶III起始前复合物的结构
2023-05-14 iNature iNature 发表于上海

该研究揭示了U6启动子上SNAPc结合的Pol III PIC的分子组织以及Pol III诱导的自发启动子开放的机制。该研究为理解SNAPc在不同复合物中对启动子的识别提供了一定结构基础。
真核RNA聚合酶III(Pol III)是转录大量短的非编码RNA所必需的,包括tRNA、5S rRNA和U6 snRNA。这些产物在各种细胞过程中发挥重要作用,如翻译、RNA加工和转录调节。
2023年5月10日,复旦大学徐彦辉及Wang Qianmin共同通讯在Cell Research在线发表题为“Structure of the SNAPc-bound RNA polymerase III preinitiation complex”的研究论文,该研究揭示了U6启动子上SNAPc结合的Pol III PIC的分子组织以及Pol III诱导的自发启动子开放的机制。该研究为理解SNAPc在不同复合物中对启动子的识别提供了一定结构基础。

在转录起始的早期阶段,不同类型的Pol III特异性基因被不同的多亚基转录因子识别,随后Pol III被募集以组装起始前复合物(PIC)。作为Pol III转录基因的原型,U6 snRNA基因包含一个TATA盒和一个近端序列元件(PSE),它们分别被TFIIIB和小核RNA激活蛋白复合物(SNAPc)识别。先前研究已经确定了PSE识别所需的SNAPc结构域或区域。尽管对Pol III延伸复合物和酵母Pol III PIC组装进行了广泛的研究,但仍不完全清楚含有SNAPc的Pol III的PIC是如何在U6启动子上组织的,以及启动子DNA是如何在PIC复合物中熔化的。
在报道的缺乏SNAPc复合物的酵母Pol III CC的结构中,由于高度的灵活性,-1下游的启动子是不可见的,并且观察到的下游启动子的末端位于远离聚合酶核心的位置。与人类CC1复合物相比,酵母CC具有更窄的裂缝,这将与人类CC2的下游DNA产生空间冲突。这可能反映了两种复合物之间的物种特异性构象差异。
该研究的冷冻电镜图支持SNAPc亚基SNAPC1、SNAPC3和SNAPC4的建模,它们形成了先前定义的mini-SNAPc8。没有观察到SNAPC5,可能是因为它是柔性的,与其在DNA结合中可有可无的作用一致。SNAPC3是与SNAPC1和SNAPC4相互作用的主要支架。SNAPC3的C末端结构域,以前称为“SNAPc指状结构域”,被SNAPC4的连接螺旋包围。这一观察结果与之前提出的黑腹果蝇SNAPC4和SNAPC3的“SNAPc指结构域”之间的相互作用一致。
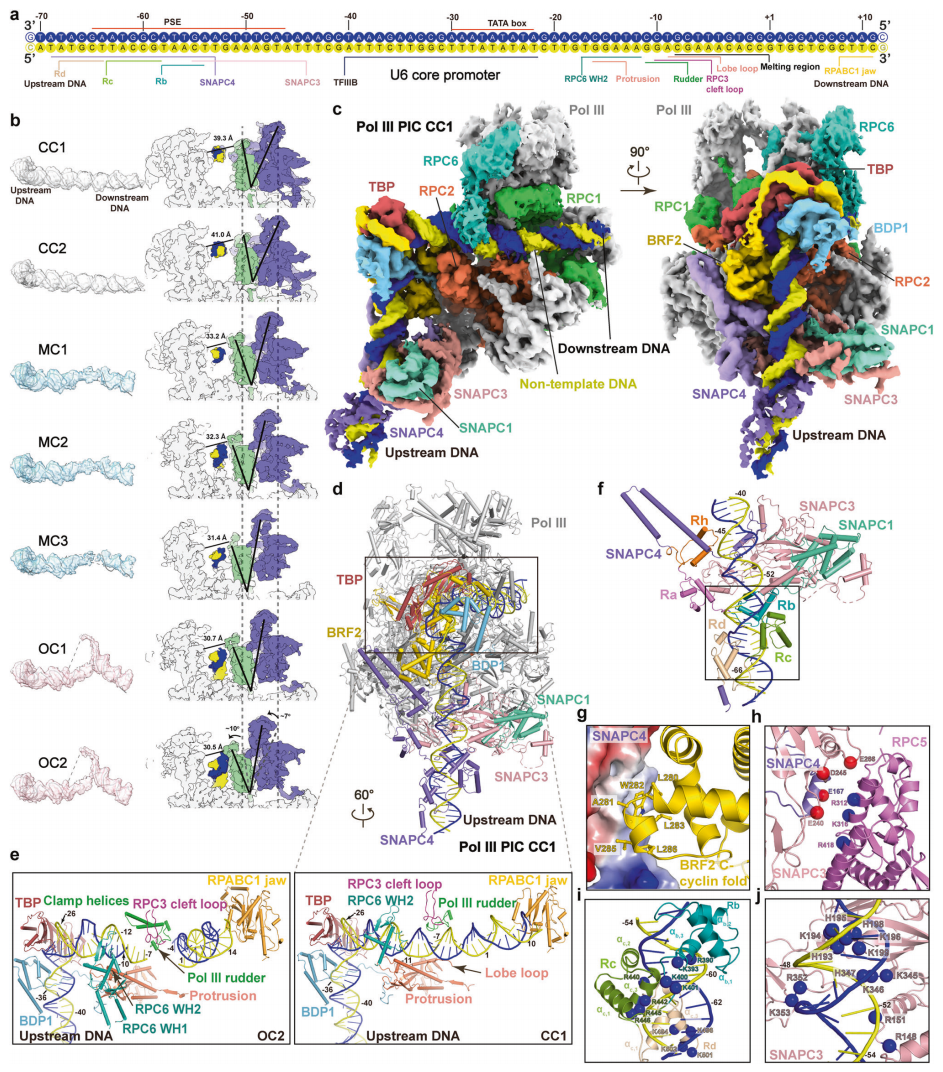
SNAPc结合的RNA Pol III PIC的冷冻电镜结构(图源自Cell Research )
TATA盒和上游PSE区主要被TFIIIB和SNAPc识别,分别产生约2400Å和约2500Å的DNA结合区。SNAPC4包含四个半MYB重复序列,称为Ra–Rd和Rh,其中Ra和Rh重复序列主要结合SNAPC3,而Rb、Rc和Rd重复序列结合启动子DNA。每个MYB重复序列由一个基本的N末端环组成,后面是三个连续的α-螺旋(α1、α2和α3),如在典型的MYB重复中所观察到的。这一观察结果表明SNAPC4的MYB重复序列是启动子结合所必需的,Rc和Rd重复序列是PSE元件识别所不可或缺的。SNAPC3通过两个高度保守的基本环从-48到-52与PSE直接接触,与SNAPC3的“SNAPc指状结构域”对启动子结合的要求一致。总之,SNAPc与PSE区进行多次接触,与TFIIIB和Pol III缔合,并可能通过组织和/或稳定PIC复合物来促进启动子打开。
综上,该研究揭示了U6启动子上SNAPc结合的Pol III PIC的分子组织以及Pol III诱导的自发启动子开放的机制。该研究为理解SNAPc在不同复合物中对启动子的识别提供了一定结构基础。
原文链接:
https://www.nature.com/articles/s41422-023-00819-x
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言